وروجکان مبتکر
شما دانش اموزان عزیز دراین وبلاگ با علوم تجربی نهم آشنا خواهید شدوروجکان مبتکر
شما دانش اموزان عزیز دراین وبلاگ با علوم تجربی نهم آشنا خواهید شددرباره من
 سلام به وبلاگ ما دو تا رفیق خوش اومدین
من و زهرا در مدرسه ی ازاده مشغول به تحصیل هستیم .
امیدواریم ازمطالبی که برای شما به اشتراک میذاریم کمال استفاده رو ببرین.
ادامه...
سلام به وبلاگ ما دو تا رفیق خوش اومدین
من و زهرا در مدرسه ی ازاده مشغول به تحصیل هستیم .
امیدواریم ازمطالبی که برای شما به اشتراک میذاریم کمال استفاده رو ببرین.
ادامه...
فصل 12

تغییراتی که در عناصر غربالی صورت میگیرد تا نهایتا به
یک عنصر فعال در انتقال تبدیل شود بسیار چشمگیر است. این تغییرات در دیوارهها ، سیتوپلاسم و اندامکها میباشد. دیوارههای جانبی سلولها زیاد تغییر نمیکنند
و فقط کمی ضخیم میشوند. در زیر میکروسکوپ با یک منظره شفاف دیده میشوند که این حالت شفاف احتمالا به
ترکیب دیواره سلولی در این عناصر بستگی دارد. مهمترین تغییرات در دیواره انتهایی
آنها صورت میگیرد که به شکل سوراخهای متعدد در این دیوارهها منجر میشوند که در
نهایت حالت غربال به خود میگیرد که آنها را به نام صفحه
آبکش میگویند.
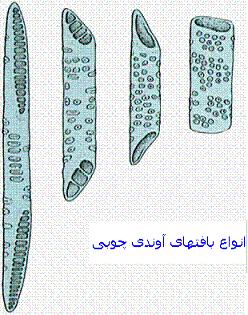
انواع صفحه آبکش
اگر صفحه آبکش سوراخهایش در همه قسمتها یکنواخت پخش شود صفحه آبکش ساده ولی اگر در قسمتهایی متراکم باشد، صفحه آبکش مرکب نامیده میشود. قطر منافذ میتواند از چند نانومتر تا چند میکرومتر تغییر کند. اکثرا حدود 1.18 میکرومتر است. ولی تغییرات آن را تا حدود 14 میکرومتر نیز گزارش کردهاند. مثلا در کدو 10 میکرومتر است.
تغییرات درون سلولی عناصر آبکش
از
جمله تغییراتی که در سیتوپلاسم عناصر آبکشی اتفاق می افتد، مهمترینشان در هسته و واکوئل است. هسته در سلولهای آبکشی در جریان تمایزیابی به تدریج
ناپدید میشود. این سلولها در ابتدای تمایز دارای واکوئلهای کوچکی هستند که به
تدریج به یکدیگر ملحق شده و بزرگ میشوند و تشکیل یک واکوئل درشت مرکزی میدهند.
در سلول آبکشی تشکیل واکوئل نشانه تکمیل تمایز نیست. بلکه در مرحله بعدی غشای
واکوئل (تونوپلاست) از بین میرود و محتویات واکوئل در سیتوپلاسم آزاد میشود. میکروفلامنتها ، میکروتوبولها ، دستگاه گلژی و ریبوزومها در سلولهای آبکشی بالغ دیده نمیشوند. اندامکهایی نظیر میتوکندری ، پلاستیدها و شبکه
یکنواخت آندوپلاسمی در سلولهای
آبکشی دیده میشود.
|
|
طول عمر سلول آبکشی
دوره زندگی سلولهای آبکشی چند هفته تا چند ماه است. در بعضی از گونهها سلولهای آبکشی در تمام یک دوره رشد و حتی در بعضی گیاهان مانند مو و زیرفون مدت دو سال زنده میمانند. در پایان زندگی سیتوپلاسم این سلولها مادهای به نام کالوز ترشح میکنند و منافذ دیواره عرضی را مسدود میسازند. با بسته شدن آوندهای آبکش عمل آنها در هدایت شیره پرورده متوقف میشود. ولی ذخیره مواد در داخل آنها میتواند ادامه پیدا کند. در فصل بعد حتی در صورت حل شدن کالوز سلولهای آبکشی میمیرند و از هوا پر میشوند. در این موقع نقش آنها در هدایت شیره پرورده ، بطور نهایی پایان مییابد.
سلولهای همراه بافت فلوئم
در
نهاندانگان در کنار هر سلول آبکشی به تعداد 1یا 2 و گاهی 3 عدد دیده میشود. این
سلولها در طول و یک طرف هر یک از سلولهای آبکشی دیده میشوند. این سلولها دارای
منشا مشترکی با سلولهای آبکشی هستند. سلول مادر آبکش با یک تقسیم نامساوی ، دو
سلول بوجود میآورد که سلول بزرگتر بتدریج تمایز پیدا کرده و به سلول آبکشی تبدیل
میشود. سلول کوچکتر منشا تشکیل سلول همراه میگردد. سلول همراه در نهاندانگان توسط پلاسموسمهای متعددی با سلولهای آبکش ارتباط برقرار میکند.
برسیها نشان داده وقتی سلول آبکش میمیرد ، سلول همراه نیز بطور همزمان میمیرد.
گاهی نیز مشاهده شده که با مردن سلول آبکش ، سلول همراه
منشا تشکیل یک سلول آبکش دیگر میگردد. از نظر سیتولوژیکی سلولهای همراه دارای
سیتوپلاسم متراکم و واکوئلهای کوچک و فراوان و هسته بزگ مرکزیاند و هرگز نشاسته در خود ذخیره نمیکنند. عقیده بر این است که سلولهای همراه
انجام برخی از اعمال متابولیکی مانند ساخت پروتئین را که در جریان تمایز عناصر
غربالی کاهش یافته یا حذف شده است بر عهده دارد. شاید میتوکندریهای فراوان موجود
در سلول همراه تامین کننده انرژی عناصر غربالی (به صورت ATP) باشد.
در برخی گونهها مواد فتوسنتزی تولید شده در سلولهای مزوفیل از طریق سلولهای همراه به عناصر
غربالی راه مییابد. در بازدانگان گاهی در کنار سلولهای آبکشی یک سلول کوچک دیده میشود که در
قسمتی از سلول آبکشی قرار دارد نه در تمام طول آن. این سلول کوچک به خاطر مکان ،
موقعیت و اختصاصات سیتولوژیکی کاملا قابل مقایسه با یک سلول همراه است و به نام
سلول آلبومینی خوانده میشود. منشا سلول آلبومینی برخلاف سلول همراه یکی از
سلولهای پارانشیمی بافت آبکشی است.
سلولهای پارانشیمی بافت فلوئم
این
سلولها از نوع ذخیرهای هستند که میتوانند ترکیباتی مانند نشاسته ، لیپیدها ، تاننها یا نشاستهها را در خود جمع
کنند. سلولهای پارانشیمی در ساختمان فلوئمهای اولیه در امتداد آوندهای آبکشی و به
موازات آنها قرار گرفتهاند و به نام پارانشیم عمودی (Vertical Phloem Parenchyma) نامیده میشود
و یاختههای پارانشیمی دیگر که در جهت شعاعی کشیده شدهاند، پارانشیم افقی یا
شعاعی (Horizontal Phloem
Parenchyma) نامیده میشوند. دیواره سلولی پارانشیم در
فلوئمهای اولیه سلولزی و در ثانویه میتواند چوبی باشد. یاختههای پارانشیمی بافت فلوئم در
اندامهای هوایی گیاهان خصوصا در شرایطی که بافت
مخاط پریدرم توسعه قابل توجهی در اندام نداشته باشد،
میتواند حاوی کلروپلاست باشد.
|
|
سلولهای فیبر بافت فلوئم
سلولهای فیبر هم فلوئم اولیه و هم در فلوئم ثانویه
دیده میشوند. این سلولها در ابتدای تشکیل خود دارای اندازه کوچکی هستند و به
تدریج در جریان رشد اندام و تمایز بافت بر قطر و خصوصا طول آنها افزوده میشود.
رشد آنها به گونهای است که سلولهای کشیده با دو انتهای باریک حاصل میشود که
منظره دوکی شکل دارد. دیواره
سلولهای فیبر از جنس لیگنین میباشد.
البته باید توجه کرد که بعضی از فیبرهای فلوئمی دیوارههایشان
برای همیشه سلولزی باقی میماند که آنها را فیبر سلولزی مینامند. فیبر کتان از این نوع
است. گاهی رسوب لیگنین در دیوارهها به قدری زیاد است که 90درصد حجم دیواره را اشغال میکند. در
این موقع سلول میمیرد. گاهی دیواره سلول فیبر دارای لیگنین کمتری هستند. اینها
معمولا میتوانند سیتوپلاسم خود را حفظ کنند و توسط پلاسمودسماتا با سلولهای مجاور ارتباط
برقرار کنند. گاهی ممکن است سلول فیبر تقسیم شود و داخل
سلول خانه خانه دیده شود . گاهی ممکن است دارای هستههای متعدد باشد.
آوند چوبی
در اکثر گیاهان محسوب میشود آوند چوبی طولانیترین بخش مسیر انتقال آب. در یک گیاه یک متری بیشتر از 99 درصد مسیر انتقال آب از درون گیاه در داخل آوند چوبی میباشد. سلولهای هدایت کننده در آوند چوبی از ویژگی خاصی برخوردارند که باعث میشود بتوانند مقادیر زیادی آب را بطور کارآمد از خود انتقال دهند. دو نوع از عناصر تراکئیدی مهم در آوندهای چوبی وجود دارند که یکی از آنها تراکئیدها و دیگری عناصر آوندی یا وسلها میباشند.
هنگامی که این عناصر فعالیت میکنند به صورت بافت بوده و
غشا یا اندامکی ندارند. بنابراین اینها مانند لولههای توخالی
هستند که دیوارههای
ثانویه چوبی به آنها استحکام میبخشد. فقدان غشای
سلولی در عناصر تراکئیدی و وجود منافذ در دیواره وسلها باعث میشود که آب در واکنش
به شیب مکش یا فشار به راحتی از این لولههای مویین عبور کند.
|
|
تراکئیدها
سلولهایی
کشیده با دو انتهای باریک هستند که در ساختمان اندامهای مختلف گیاهی به صورتهای
خاصی پشت سر هم قرار میگیرند طوری که انتهای باریک آنها به صورت همپوشان واقع میشود.
یاختههای تراکئیدی با این آرایش منظره خانه خانه به خود میگیرند که با نام آوند
ناقص خوانده میشوند. در جریان تمایزیابی تراکئیدها سلولها به تدریج محتویاتشان را
از دست میدهند و در پایان تمایزیابی میمیرند و به شکل محفظههایی توخالی درمیآیند.
پدیده تمایزیابی در این بافتها با لیگنینی شدن دیواره
همراه است. ماده لیگنین پیش از مرگ سلولها به صورت ناپیوسته و به اشکال
مختلف روی دیواره اولیه نهاده میشود. تزئینات لیگنین که به این ترتیب تشکیل میشود.
بر اساس نحوه رسوب لیگنین انواع مختلف تراکئید مثل حلقوی و مارپیچی را در نهانزادان
آوندی و هالهای یا قرصی را در بازدانگان بوجود میآورد. سلولهای تراکئیدی به علت نحوه قرارگیری در
اندامهای گیاهی و داشتن لیگنین نه تنها در نقش انتقال دهنده در پیکر گیاه میباشد
بلکه نقش استحکام بخش را نیز بر عهده دارد. انتقال آب و املاح معدنی در تراکئیدها
از طریق لانها یا پیت صورت میگیرد.
وسلها
فقط
در نهاندانگان دیده میشود و به تعداد متغیر در ساختمان یک آوند شرکت میکنند
هر کدام از این یاختهها را به عنوان یک عضو تشکیل دهنده وسل گویند. آوندهای که به
اینگونه ساخته میشوند آوند کامل نامیده میشوند. تمایزیابی در هر کدام از سلولهای
وسل به گونهای است که نهایتا منجر به مرگ سلولها میشود. همچنین در مراحل پایانی
تمایز دیواره عرضی یاختهها هم از بین میروند.
در محل دیوارههایی که در طی تمایز متلاشی میشوند در
بسیاری از موارد یک حفره بزرگ ایجاد میشود. در پایان تمایز دیواره انتهایی کاملا
از بین میرود. در بعضی از گیاهان دیواره انتهایی از چند سوراخ بزرگ تشکیل مییابد
نه یک حفره بزرگ و ندرتا دیده میشود که حالت مشبک دارد. به دنبال از بین رفتن
دیواره انتهایی ، آوندها تشکیل میشوند که تعداد سلولهای تشکیل دهنده آنها متفاوت
است.
سلولهای پارانشیمی بافت گزیلم
سلولهای
پارانشیمی بافت گزیلم همانند بافت
فلوئم از نوع ذخیرهای اند. این سلولها دارای بافت سلولزی یا لیگنینی میباشند. خصوصا در گزیلم اولیه ، پارانشیمهای
گزیلمی به گونهای هستند که در امتداد عناصر هدایت کننده این بافت قرار میگیرند.
ولی در گزیلمهای ثانویه علاوه بر پارانشیمهای عمودی ، پارانشیمهای شعاعی در امتداد
شعاعی مشاهده میشود.
سلولهای پارانشیمی چه پارانشیم شعاعی و چه پارانشیم
عمودی معمولا تا زمانی که عناصر هدایت کننده بافت گزیلم دارای فعالیت انتقالی میباشند،
زنده هستند زمانی که این اعضا غیر فعال شوند، پارانشیمها نیز میمیرند. ولی مواردی
ممکن است یاختههای پارانشیمی تا چند سال پس از غیر فعال شدن آوند زنده باشند.
گاها دیده میشود که سلولهای پارانشیمی دورتادور آوند به صورت خاصی قرار میگیرند.
این سلولها همه اطراف آوند را مانند غلاف فرا میگیرد در چنین شرایطی به آنها Vessel associated paranchym گویند.
|
|
انواع تغییرات دیواره سلولهای پارانشیمی
سمتی از سلولهای پارانشیمی که به طرف گزیلم قرار دارد در محل دیوارههای سلولزی خود ، تغییرات قابل توجهی را نشان میدهد این سلولها به طرف داخل سلول برجستگیهایی را ایجاد میکنند که به شدت توسعه مییابد و نتیجه آن افزایش سطح غشاست. این غشا نقش مهمی میتوانند در انتقال مواد داشته باشند. و آنها را Transfer cellsیا یاختههای ناقل مینامند. این نوع گیاهان را بطور خاص در گیاهان علفی در محل گرهها و برگها و در برخی از ساختارهای ترشحی مشاهده می کنیم
.
زمانی که سلولهای هدایت کننده بافت گزیلم به خاطر افزایش
سن گیاه غیر فعال شود یا اینکه غیر فعال شدن آوندها به دنبال ایجاد جراحت یا وارد
آمدن هر نوع آسیب به بافت اتفاق افتد، ملاحظه میشود که سلولهای پارانشیمی مجاور
زوائد پروتوپلاسمی متعدد ایجاد میکنند که از طریق پونکتاسیونهای موجود میان یاخته
پارانشیمی و وسلها به داخل آنها نفوذ میکند. تعداد این زوائد میتواند زیاد باشد
ضمنا این زوائد داخل آوند بزرگ و بزرگتر میشود و نهایتا آوند را مسدود میکند .
این زوائد سیتوپلاسمی تیلوز خوانده میشود.
با بسته شدن آوند ، پاتوژنها و انگلها نمیتوانند به درون آوند نفوذ کنند.
سلولهای فیبری بافت گزیلم
فیبرها در گزیلمهای اولیه و ثانویه دیده میشوند. سلولهای فیبر در بافت گزیلم دارای دیوارههای لیگنینی هستند. ولی ضخامت لیگنین در دیواره سلولها برحسب انواع مختلف آنها متفاوت است. انواع فیبر در بافت گزیلم عبارت است از :
فیبر تراکئیدی
در این نوع فیبر ، ضخامت ماده لیگنین در دیوارههای سلولی بسیار اندک است. این نوع فیبرها معمولا زنده هستند و بوسیله پلاسمودسماتا با سلولهای مجاور در ارتباط اند.
فیبر لیبری فرم
این
فیبرها شبیه به فیبرهایی هستند که در ساختمان
فلوئم ثانویه مشاهده میشوند. اصطلاح Liber به معنی
فلوئم ثانویه است و این فیبرها را به علت تشابه شان به فیبرهای موجود در لیبر ،
لیبری فرم گویند. این فیبرها دیواره ضخیم لیگنین دارند که حجم اعظم دیواره را
اشغال میکند و اغلب سلولها مردهاند.
|
|
فیبر خانه خانه
اینها فیبرهایی هستند که دارای دیواره عرضی متعدد میباشند. به این صورت که سلول فیبر با تشکیل دیوارههای افقی چند بار تقسیم میشوند و در نتیجه فیبر یک منظره خانه خانه بخود میگیرد (در برش طولی). این فیبرها اغلب میتوانند ترکیبات شیمیایی با ماهیتهای مختلف را در خود انباشته کنند. این مواد بیشتر ترکیبات نشاستهای ، لیپید ، تانن ، و ترکیباتی مثل اگزالواستیک اسید است.
فیبر ژلاتینی
فیبرهای ژلاتینی نوع دیگری از فیبرها هستند که در ساختمان چوبی به نام چوب واکنش تشکیل میشوند. این نوع فیبر در ساختمان چوب نهاندانگان در شاخههایی که حالت افقی یا مورب دارند، دیده میشود. این نوع فیبرها یا ماده لیگنینی ندارند یا لیگنین آنها کم است. ترکیبات پکتیکی در این نوع فیبرها فراوان است .
مقایسه کاج و سرو
دو درخت کاج و سرو در ظاهر شباهت زیادی به هم دارند اما در حقیقت از نظر ویژگی ها باهم متفاوت اند .


1 درخت کاج
این درخت به صورت قامت بلند و
با شیارهای طولی روی تنه که تاج آن به صورت شاخه های عمودی و افقی رشد کرده با سوزن
های جدا از هم و بلند می باشد و به علت فاصله کم کاشت این درختان در خیابان ها و
رشد آپارتمان سازی در شهر بزرگی مثل تهران ، باعث شده که رشدشان به خطر افتد و به
علت بلندی ساختمان ها و نرسیدن نور کافی ، گاهاص در خیابان ها و کوچه ها این
درختان را با تاج و تنه کج ببینیم .
چنانچه شاخه های جانبی (روی تنه ) و یا انتهایی (نوک
درخت ) را قطع کنیم به هیچ وجه جایگزین نخواهند شد و به علت رشد کند محل این شاخه
های قطع شده لخت خواهد ماند، پس هرگز نباید سربرداری و یا هرس روی این درختان
انجام شود، چون به علت عدم جایگزینی شاخه های جانبی و طولی فرم درخت شکل نامناسبی
پیدا می کند. میوه این درختان بزرگ و مخروطی شکل می باشد و شاید بتوان آن را شبیه
یک نارنجک دستی دانست و تاج این درختان محل مناسبی برای زندگی و لانه سازی کلاغ ها
بوده که همه ما در شهرها و یا همه جا که این درختان کاشته شد، مشاهده کردیم . ریزش
سوزن و میوه این درختان زیاد بوده و از این نظر مشکلات زیادی را از نظر نظافت پای
این درختان به وجود می آورد.
شاید گذرمان به اتوبان چمران و
یا خیابان فاطمی و یا بهشت زهرای تهران و جنگل سراوان جاده رشت و قزوین افتاده
باشد، آن وقت می توانیم تعداد زیادی از این درختان را با قامت های بلند و کوتاه
ببینیم .
2درخت سرو
این درختان اکثرا در حاشیه
شهرها و کمربند سبز تهران و پارک های شهری به صورت انبوه کشت شده و دارای سوزنهای
به هم پیوسته و گوشتی و ریز می باشد.
این درختان با توجه به اینکه
دارای قدرت بالایی از جهت سبز شدن روی شاخه های جانبی و طولی بوده همیشه با پوشش
سبز در سرتاسر تنه دیده می شود و با انجام هرس روی این درختان شکل های زیبایی را
می توان به وجود آورد درختان کهنسال سرو دارای میوه های گرد و کوچک است که در بعضی
گونه ها به بزرگی گردو نیز دیده می شود. ریزش سوزن و میوه در این درختان بسیار کم
می باشد.
به عنوان مثال دو گونه را که
زیاد در شهرها و ارتفاعات به چشم مان می خورد نام می برم .
سرو شیراز: که دارای قامت بلند
بوده و شاخه های جانبی آن بطور عمودی رشد می کند و تمام تنه را می پوشاند و با
انجام هرس روی آن می توان اشکال زیبایی را به وجود آورد که در باغ ارم شیراز و
پارک های لاله و ساعی تهران تعداد زیادی از آن را می توان دید.
یک گونه دیگر سرو نقره یی نام
دارد که به علت مقاومت زیادش به کم آبی آن را می توان در خاک های نامرغوب نیز کاشت
و به همین علت در ارتفاعات تهران و جاهایی که دسترسی به آب کم می باشد می توان به
صورت انبوه مشاهده کرد. در این درختان به علت رشد سریع و امکان هرس روی آن شکل های
مختلف زیبایی را می توان به وجود آورد، البته ما اغلب این درختان را به صورت
مخروطی شکل و انبوه در حاشیه شهر و پارک ها مشاهده می کنیم و اگر از اتوبان مدرس و
یا آزادگان تهران عبور کرده باشیم می توانیم تعداد زیادی از این گونه را مشاهده
کنیم . بطور ساده درختان کاج دارای قامت بلند با تنه قهوه یی با شیارهای طولی و
لخت هستند و دارای سوزن های بلند و جدا از هم بوده و قابل هرس نیستند. ولی درخت
سرو دارای تنه کاملاص سبز و پوشیده از سوزن های ریز گوشتی به هم چسبیده (مرکب )
بوده و دارای قامت کوتاه تری نسبت به درختان کاج می باشد و می توان روی این درختان
عملیات هرس انجام داد و اشکال مختلف و زیبا به وجود آورد.
چرخه زندگی سرخس
یکی از گیاهان زیبای آپارتمانی که جنبه زینتی دارد سرخس است این فوژر آمریکایی استوایی را باید یکی از جالب ترین گیاهان دانست. برگ های دراز و هلالی شکل آن از قلب گیاه می روید و با حالت پرمانند و انبوه خود چه در باغچه و یا روی میز کنسول ها و کنار مجسمه ها جنبه تزئینی بسیار زیبایی دارد...

درپشت برگ شاخه ها لکه های قهوه ای مایل به نارنجی موسوم به هاگینه مشاهده می شود. هرهاگینه مجموعه ای از هاگدان های گرز مانند است.
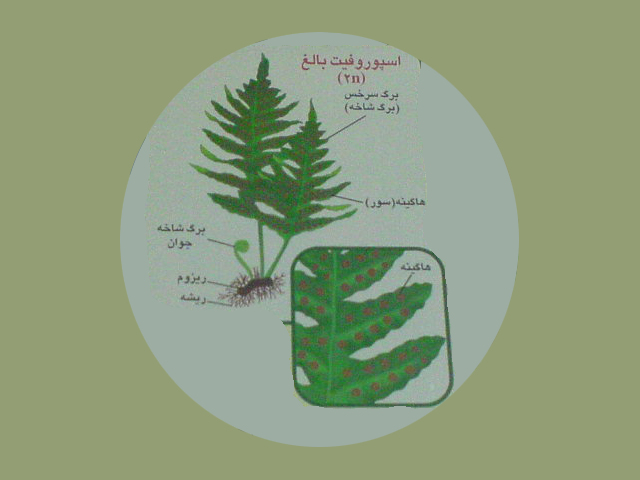
درون هاگدان سلول های مادر هاگ دیپلوئید،باتقسیم میوز هاگ های هاپلوئید را به وجود می آورد. پاره شدن هاگدان ها منجربه آزاد شدن هاگ ها می شود.

ازرویش هاگ گامتوفیت تشکیل می شود
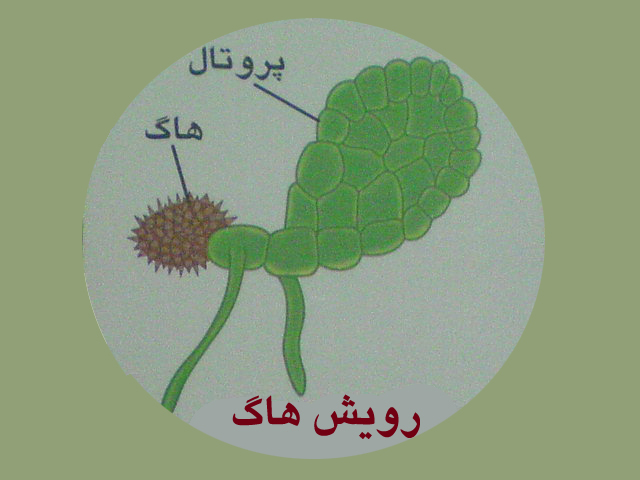
گامتوفیت سرخس صفحه ای سبزرنگ وقلبی شکل است و پروتال نام دارد. پروتال زندگی مستقلی دارد . درسطح زیرین پروتال ابتدا آنتریدی سپس آرکگن تشکیل می شود .
آنتریدی تولیدآنتروزوئیدمی کند ودرون آرکگن تخمزا تشکیل می شود.
آنتروزوئیدهای سرخس که تاژک های متعدددارنددرآب شنا کرده به طرف آرکگن ها حرکت می کنند، وارد آرکگن شده وباتخمزاترکیب می شوند ، حاصل لقاح آنتروزوئید وتخمزا ، سلول تخم است .
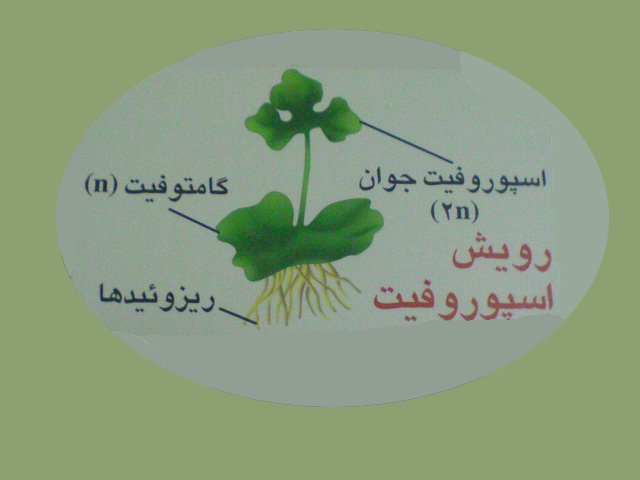
ازرویش سلول تخم اسپوروفیت جوان تشکیل می شود . اسپوروفیت سرخس فقط در ابتدای رویش به گامتوفیت وابسته است ، سپس مستقل شده ، رشد نموده وبالغ می شود .
چرخه زندگی کاج
نوعی درخت از دسته مخروطیان است که برگهای دائمی دارد. کاجها به ترتیب در شاخههای دانهداران، بازدانگان، مخروطداران قرار دارند.
پولکهای مخروط کاج همان برگهای کاج هستند که تغییر شکل یافتند.

اندام های تولید مثلی کاج در مخروط ها تشکیل می شود . کاج دونوع مخروط دارد ، مخروط نر ومخروط ماده .


مخروط ماده شامل محورطولی وپولک هاست . در سطح بالایی هر پولک دو تخمک تشکیل می شود . مخروط نر نیز شامل محور طولی وپولک هامی باشد . درسطح زیرین هر فلس دوکیسه گرده تشکیل می شود .
درکیسه های گرده ، سلول های مادرمیکروسپور دیپلوئید باتقسیم میوز تقسیم شده ، چهار میکروسپور هاپلوئید رابه وجود می آورد .
هسته هرمیکروسپور دومیتوزمتوالی انجام داده چهارهسته تشکیل می شود .هرهسته تبدیل به یک سلول می شود .
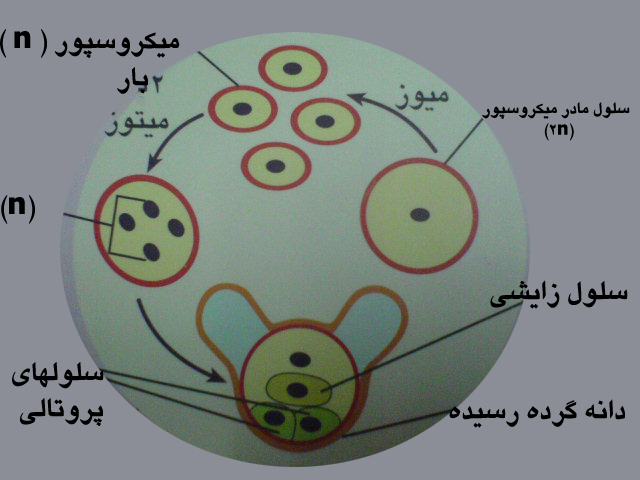
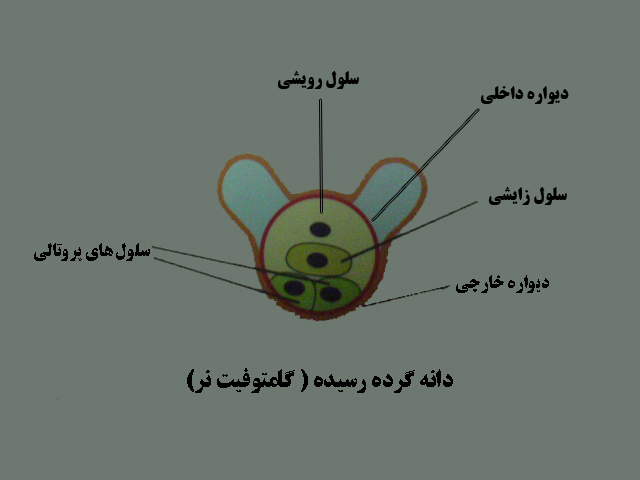
به این ترتیب دانه گرده کامل می شود . میکروسپور یا دانه گرده همان گامتوفیت نر است . هردانه گرده رسیده شامل چهار سلول هاپلوئید است که عبارتند از یک سلول زایشی ، یک سلول رویشی ودوسلول پروتالی که دودیواره خارجی وداخلی آنهارا در احاطه می کند. دیواره خارجی در دوطرف مقداری از دیواره داخلی فاصله می گیرد ودوبال را ایجاد می کند .
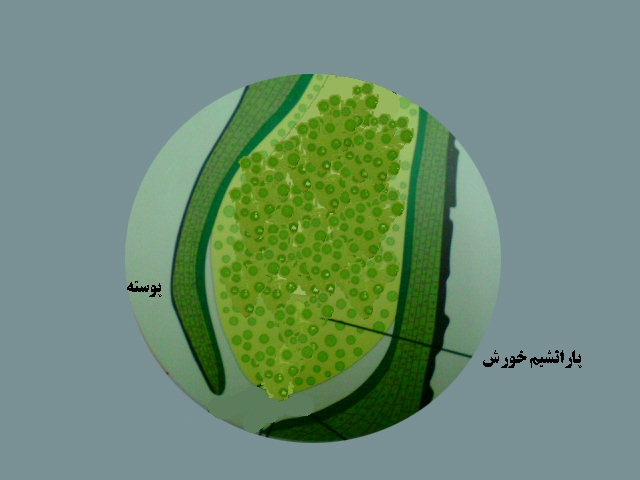
پس ازتشکیل دانه گرده گرده افشانی انجام می شود . تخمک ها درسال اول نارسند . تخمک نارس کاج شامل یک پوسته ، سفت وپارانشیم خورش است .
در سال دوم تخمک ها می رسندبه این ترتیب که یکی از سلول های پارانشیم خورش به نام سلول مادر مگاسپور به روش میوز چهار مگاسپور را به وجود می آورد . سه تا از این سلول ها ازبین رفته ، مگاسپور باقی مانده با میتوز های متوالی یک بافت هاپلوئید به نام آندوسپرم را تشکیل می دهد . آندوسپرم همان گامتوفیت ماده است . درون آندوسپرم تعدادی آرکگن ودرون آرکگن ، تخمزا تشکیل می شود .

درطول این مدت براثر فعالیت سلول رویشی ، لوله گرده تشکیل شده که ازسفت عبور کرده وارد تخمک می شود .پس از تشکیل لوله گرده ، سلول رویشی تحلیل می رود. سلول زایشی با تقسیم میتوز دو آنتروزوئید را به وجود می آورد . آنتروزوئیدها از طریق لوله گرده وارد تخمک شده وبا تخمزای درون آرکگن ترکیب می شوند. به این ترتیب تخم تشکیل می شود . از رشد سلول تخم، رویان تشکیل می شود . رویان همان اسپوروفیت آینده است .

پس از لقاح ، تخمک تغییر کرده تبدیل به دانه می شود .دانه کاج شامل پوسته ، رویان وآندوسپرم است که اندوخته غذایی دانه را تشکیل می دهد. دانه کاج یک بال نیزدارد که به پراکنده شدن دانه کمک می کند .
درشرایط مساعد دانه می روید واسپوروفیت جدیدی را ایجاد می کند.

می توان گفت که اسپوروفیت کاج فقط در ابتدای رویش دانه به گامتوفیت وابسته است زیرا از آندوسپرم تغذیه می کند.
چرخه زندگی خزه
خزه ها گیاهانی اند که از روش تولید مثل غیر جنسی قطعه قطعه شدن تولید مثل می کنند ...

گامتوفیت ( n) گیاه اصلی وشناخته شده است.
گامتوفیت شامل ضمائم برگ مانند ، محور ساقه مانند و ریزوئید است .

ازرویش هاگ ها دونوع گامتوفیت تولید می شود



اندام تولید مثل نر : آنتریدی
اندام تولید مثل ماده: آرکگن
آنتریدی باتقسیم میتوز آنتروزوئیدهارابه وجود می آورد ودرون آرکگن تخمزا تشکیل می شود.
آنتروزوئیدهای دوتاژکی خزه درآب شنا کرده وارد آرکگن می شوند، حاصل لقاح آنتروزوئید وتخمزا ،سلول تخم است . سلول تخم درون آرکگن رشد نموده، اسپوروفیت جوان راتشکیل می دهد.

اسپوروفیت برروی گامتوفیت ماده تشکیل واز آن استفاده می کندبه همین دلیل گفته می شود اسپوروفیت ،انگل گامتوفیت است .



اسپوروفیت جوان رشدکرده وبالغ می شود. اسپوروفیت بالغ شامل سه بخش پایه ، تار وکپسول است . کپسول همان هاگدان است . درون هاگدان باتقسیم میوز هاگ های هاپلوئید به وجود می آید
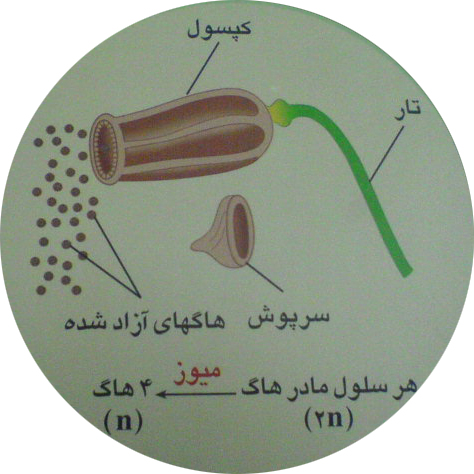
ازرویش هاگ گامتوفیت های جدید تشکیل می شود به این ترتیب چرخه زندگی خزه تکرار می شود.
گیاهانی که کاغذ می شوند
برای دانلود pdf گیاهانی که کاغذ می شوند اینجا کلیک کنید.